|
|||||||||||||
| Home | Research | People | Publications | BBS | Photos | Contact Us | |||||||
|
4. Conservation genetics with emphasis on the rare and endangered plants The development of conservation biology in China has been relatively slow for many years due to historic reasons, and less emphasis has been focused on genetic issues in researches and management of biodiversity. During recent decades, we have used different molecular markers to perform preliminary population genetics investigations on many rare and endangered species, including wild rice (e.g., Ge et al. 1999 PNAS; Zhou et al. 2003 TAG; Gao et al. 2005 Biodivers Conserv; Qian et al. 2005 Genet Resour Crop Ev; Zhou et al. 2008 TAG), conifers (Ge et al. 1998 IJPS; Wang & Ge 2006 ME; Tian et al. 2008 AB), orchids (e.g., Sun et al. 2006 Bot J Linn Soc; Sun et al. 2009 AB; Li & Ge. 2006 PSE), and many other wild species (e.g., Ge et al. 1997 Conservation Biology; Bao et al. 2005 Genet Resour Crop Ev; Lopez-Pujol et al. 2006 Biodivers Conserv; Lopez-Pujol et al. 2008 Biochem Genet; Sun et al. 2006 Biochem Genet; Xu et al. 2008 TAG). In a molecular phylogeographic study of Cathaya argyrophylla, an endangered conifer endemic to China, we used sequences of maternally inherited mtDNA and biparentally inherited nuclear DNA to have found significantly lower nucleotide diversity within populations but significantly higher divergence among populations/regions in this endangered species relative to other conifers (Wang & Ge 2006 ME). In conjunction with previous allozyme and RAPD analyses (Ge et al. 1998 IJPS), we demonstrated that extant populations of C. argyrophylla may have been derived from four refugia during the Quaternary glaciations, and their populations have not expanded afterwards (Figure 4-1). The great decrease of population sizes due to the glaciations could result in strong bottleneck effects in the species, which would further lead to low genetic diversity, particularly at the population level, and high genetic differentiation among populations that was rarely observed in other gymnosperms. As a consequence, we recommended an ex situ conservation and artificial crossing for the management of this endangered species (Wang & Ge. 2006 ME). Recently, we have developed 50 SSR markers for this species, which would facilitate further studies on population genetics and evolutionary history of this endangered conifer (Wang et al. 2010 JSE).
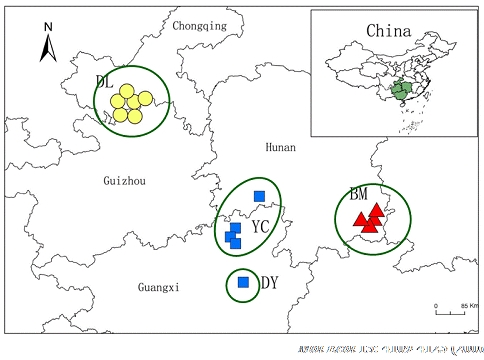
Figure 4-1. The potential refugia of C. argyrophylla during the Quaternary. Four refugia have 3 mitochondrion haplotypes (triangle, square, and circle). Changnienia amoena is an endangered orchid endemic to China. Using RAPD markers we detected low within population but high among population variation in this orchid, which is likely due to the small population sizes and local extinction because of habitat destruction (Li & Ge 2006 PSE). Based on careful examination of life history characteristics and pollination biology of C. amoena, we corrected previous recognition of the way of reproduction in the species, and found that this orchid was exclusively pollinated by two bumblebees (Bombus trifasciatus and B. imitator) (Figure 4-2). In addition, we observed that the lack of enough pollinators in C. amoena should be mainly responsible for its low fecundity and further resulted in the endangerment of the species (Sun et al. 2006 Bot J Linn Soc). A recent pollination study demonstrated that pollen transfer occurred mainly within populations and restricted gene flow by pollen and seeds probably explains the high degree of genetic differentiation between populations in this species (Sun et al. 2009 AB). These results suggest that the low within population but high among population variation in C. amoena may be attributed to the small population sizes and local extinction because of habitat destruction and loss. Therefore, we proposed that it is important to protect both the habitat and the pollinators, and recommend the artificial pollination to facilitate the restoration of the small populations (Sun et al. 2006 Bot J Linn Soc; Li & Ge. 2006 PSE).
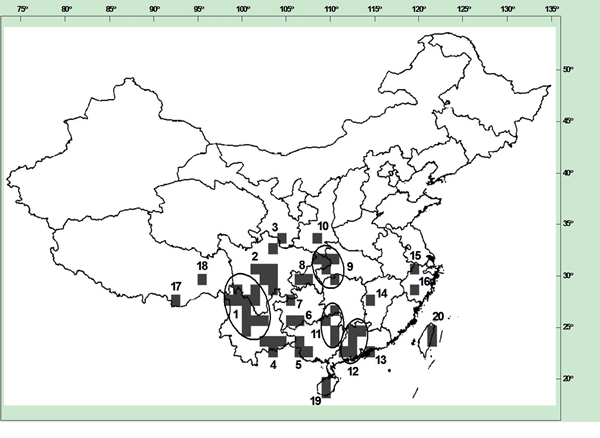
Figure 4-2. Pollinators visiting Changnienia amoena flower. A. Bombus trifasciatus probing into C. amoena. B: The pollinators, B. trifasciatus (left) and B. imitator (right). It was estimated that the rate of genetic exhaustion was much faster than that of species extinction. Thus the studies and conservation of genetic diversity is a pressing issue. China is very rich in genetic resources, but less investigation on the resource has been conducted, neither has the conservation of these resources been taken into consideration scientifically. Hereby our group is going to continue our efforts on conservation genetics studies of many species of economic values as well as the rare and endangered species. Study of this type will undoubtedly shed light on evolutionary history of plant species and facilitate further conservation management and sustainable utilization as well. By identifying the 'centres of endemism' of the Chinese spermatophyte flora, we detected the locations of past glacial refugia and assessed the role of these areas as places for plant survival ('plant museums') and/or areas for plant evolution and speciation ('plant cradles') (López-Pujol et al. 2011. J Biogeogr). We identified 20 areas of endemism in central and southern China (Figure 4-3). Although almost all centres of endemism contained both palaeoendemic and neoendemic taxa, considerable differences in their respective numbers were recorded, with the majority of neoendemics on the eastern fringe of the Tibetan Plateau (Hengduan Mountains sensu lato) but more palaeoendemics towards the east. As demonstrated in this study, owing to their varied topography, the mountainous regions of central and southern China have provided long-term stable habitats, which allowed palaeoendemics to persist and facilitated the process of speciation. Contrasting patterns between the palaeoendemics and neoendemics within refugia might be attributable to the geological and tectonic history of specific areas. The eastern fringe of the Tibetan Plateau clearly constitutes the 'evolutionary front' of China, probably as a result of the uninterrupted uplift of the plateau since the late Neogene.
Figure 4-3. The 20 centres of plant endemism in China. The shaded cells are those scoring within the top 5% highest values for weighted richness. The code numbers for the centres of endemism correspond to those in Table 1 of López-Pujol et al. (2011).
|
| © The Ge Laboratory Institute of Botany, the Chinese Academy of Sciences |